
苦草屬(Vallisneria)植物的生態學研究概況
沉水植物作為主要初級生產者,在水生生態系統中有著不可替代的作用。當沉水植物豐富時,水體表現為水質清澈、溶解氧高、藻類密度低、生物多樣性高等特點。因此,富營養化水體中沉水植物的恢復與重建已成為水域生態學研究的重點內容之一。苦草(Vallisneria)是淡水湖泊中常見的一種沉水植物。本文探討了苦草屬植物的生態學研究概況。
一、種屬與分布
1.1 種 屬
恩格勒系統(1964)
被子植物門 Angiospermae,單子葉植物綱 Monocotyledoneae,沼生目 Helobiae,水鱉亞目 Hydrocharitineae,水鱉科 Hydrocharitaceae,苦草屬 Vallisneria linn:本屬全球范圍內有10種,我國有3種,南北各省區均產。

1.2 植物特征


1.3 苦草的生物分布
苦草屬植物廣泛分布于世界各地,尤以熱帶、亞熱帶和暖溫帶多見。在我國苦草屬植物廣泛分布于南北各省區,在淡水湖泊、溝渠、池塘、內河航道的靜水或流動水體中均能很好地生長。密刺苦草是廣東江段的建群種,長勢好,蘊藏量大;而廣西江段雖然還有密刺苦草,但其優勢地位已逐步讓位給亞洲苦草(V. asia -tica) ;到了云南江段卻是苦草(V. natans)占據優勢。目前在湖北洪湖和武漢東湖也先后報道了該種的存在。
刺苦草是長江中下游湖泊的優勢物種,苦草為常見種,而密刺苦草(V. denseserrulata)僅有很少分布,苦草和刺苦草常混生于這些湖泊中,形成共存分布格局:刺苦草和苦草種間呈明顯的的帶狀相間分布格局,或苦草只零星分布于刺苦草群落中。
二、繁殖特征及生活史
2.1 生長季節及溫度
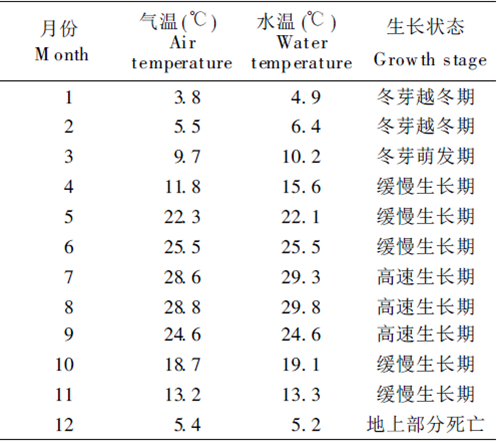
苦草是多年生草本。分布在亞熱帶地區的苦草( V. natans )冬季地上部分死亡,以冬芽或種子越冬,春季從冬芽或種子開始萌發,且隨氣溫上升開始緩慢生長,夏季為快速生長期,最大生物量一般在9月份出現。開花期是5~ 8月,結果期為8~10月,種子成熟期在12月左右。
2.2 繁殖方式
苦草的繁殖方式:分為有性繁殖(種子繁殖)和營養繁殖(冬芽和活體植株)。
1、有性繁殖
植物產生有性生殖細胞(配子),經過兩性生殖細胞的結合,成為合子,在由合子發育成為新個體(新的植株)的生殖方式,叫植物有性繁殖。
苦草具有強大的有性繁殖能力,苦草為雌雄異株,雄花在水面借助于空氣和水流擴散時可能遇到雌花,當被水波暫時淹沒時雄花可能被捕獲到雌花花被里面而完成受粉過程。
1)繁殖選擇
水深、風浪、光照、氣候環境,營養狀況對苦草對繁殖方式選擇的主要影響因素。
2)授粉
苦草分為雌株和雄株,雄株授粉完成后就會死亡,雌株多年生。水媒傳粉。
3)結果
在傳粉以后把幼果拉入水中,在水面下約30cm處比較穩定的環境里生長(25~ 30℃)。
4)萌發
萌發率為 40% ~ 69%,天然土效果最好。 萌芽率受溫度、種子保存時間和光照的影響,不受水質的影響。

2、營養繁殖
植物營養繁殖是指由植物體的根、莖、葉等營養器官或某種特殊組織產生新植株的生殖方式,屬于廣義的無性生殖范疇。
一般而言,無性繁殖方式在苦草生活史中起著更為重要的作用,冬芽和活體植株是苦草屬植物最重要的無性繁殖器官,苦草在水底地面以無性繁殖方式迅速發展可以快速占據更廣大的生境,以從基質中獲得更多的營養。
苦草的營養繁殖受以下幾種因素影響:
1、種群密度:在風浪較大的湖泊中恢復苦草種群應主要利用冬芽或鮮體,種子只適用于相對靜止的淺水池塘種植。種植必須達到一定的種群密度。經調查太湖苦草冬芽平均90~ 226個/m2。
2、光照:塊莖發芽對光照需求不大,但溫度對塊莖的發芽率有較大影響,20℃時發芽率最高,達90.2%± 12.6%,光照對幼苗生長有一定影響。苦草的塊莖苗初期的伸長生長主要依賴于基部的根莖生長。
3、種植深度:苦草球莖的萌發受土層厚度的影響很大,被5cm土層覆蓋時萌發率為100%;土層厚度為15cm時萌發率為73.3%;土層厚度為25cm時萌發率僅為3.3%。
4、機械損傷:苦草葉片生長最活躍的部位是在葉片下端2cm處,即綠色組織與白色組織的過度區,只要不破壞葉片基部的生長點,苦草的再生就非常迅速,當破壞了葉片的基部時,植株將逐漸死亡。


冬芽繁殖 活體植株
三、氮磷營養鹽循環
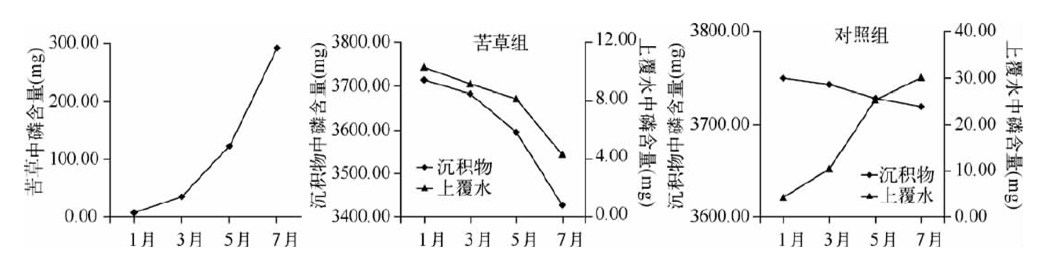
苦草對水體營鹽的去除率隨生物量的增加不斷增大,當生物量達到 2380.00g時,去除率放緩。苦草腐敗釋放的磷大部分被底泥吸附,少量進入水體。草腐敗分解具有階段性,前36d分解迅速,以后分解速率顯著下降。
3.1 尿素氮的閾值范圍
在富營養狀態下(N:4 mg/ L;P:0.2 mg/ L)不同比例硝態氮(NO-3-N)和尿素氮(Urea-N)對苦草(Vallisneria atans)的生理影響:隨著培養液中尿素氮比例的上升,生物量和硝酸還原酶活性,依次降低;POD和脲酶活性先升高后降低.研究表明,添加一定量的硝態氮可以促進苦草的生長,但當環境中尿素含量逐漸增加時會對苦草產生一定的脅迫作用,影響苦草的正常生長.從本實驗研究結果綜合來看,硝態氮(NO-3-N)和尿素氮(Urea-N)對苦草產生脅迫影響的閾值比為1∶ 1.此時TN為4 mg/ L,NO-3-N和Urea-N均為2 mg/ L。
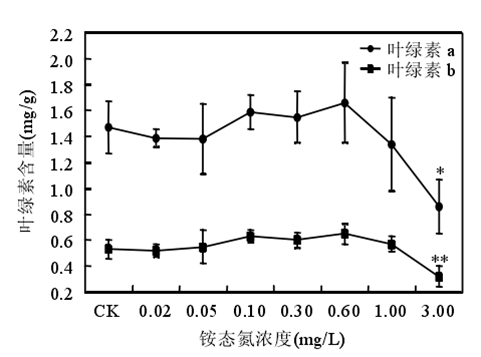

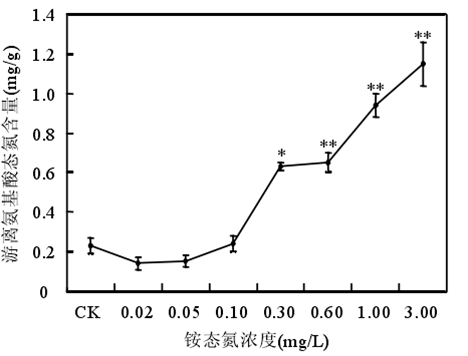
3.2 氨氮的閾值范圍
銨態氮濃度低于 0.60mg/時苦草生長良好,濃度大于或等于 1.00mg/L 時苦草的光合能力受到抑制、代謝受到干擾。苦草對銨態氮最敏感的生理生化指標是葉片中游離氨基酸態氮含量。銨態氮作為沉水植物的一種營養物質,當其含量較低時,植物由于營養缺乏誘導產生自由基。
四、種群生態特征
4.1 生物量
在很多湖泊植被調查工作中都對苦草群落或含苦草的群落生物量進行了測定(見下圖)。從表中可以看出,最高和最低生物量之間存在上千倍的差異。這一方面是因為調查對象尚包括其它種類,而非苦草本身; 另一方面,也是由于調查方法本身所致:
(1)調查結果往往用濕或鮮重表示。由于水生植物含水量豐富,離水后,因種類、天氣、采樣等因素的影響,其重量在短短的30 min內即能發生很大的變化。因此在對不同研究者的數據進行比較分析時,這便成為主要的誤差來源。
(2)各研究中取樣方法不規范,且各個方法的樣方數和重復次數過少。樣方重復次數過少常常導致結果存在極大的誤差。因為從現有的研究看,群聚分布是我國水生植物最普遍的分布式樣。
(3)測量時期不一致。一般各個研究均在最大生物量時期測量一次,但不同研究者對出現最大生物量時期看法并不一致,從6~ 9月均有,沒有逐月的調查數據是難以得出準確的最大生物量出現時期的。
從部分湖泊的工作來看, 苦草生物量還隨季節有明顯的變化,同一湖泊不同地點以及同一湖泊不同年度苦草的生物量也有較大變化,但這些工作依然或多或少存在上述的取樣問題。生物量的測定是研究植物生產力的重要基礎,關于取樣方法、樣方數量、結果表達等方面國際上已存在被普遍接受的原則,為了準確評價水生植物的生產力,同時為了減少誤差和進行國際交流,建議在今后的研究中應當以這些標準方法為指導。
(1).jpg)
4.2 群落特征
在我國多數水生植被調查工作中,群落的劃分與命名一般采用優勢度途徑,而優勢種的認定在許多情況下是以目測的表觀優勢度為標準,加之取樣方法上存在的問題,多數工作缺乏嚴格的定量分析數據。
目前研究中涉及的不同群落僅僅是具體的地方群落( Local community) ,而并非群落分類學意義上的群落類型。根據近年來的研究,我們認為層片( synusia )途徑是一個比較理想、實用的水生植物群落分類標準。利用層片分類系統,有可能對調查涉及的每一群落作出細致的分析。

五、個體生態特征
5.1 低溶解氧對苦草繁殖的影響
1、溶解氧平均值為1.6 mg/L時對苦草株重、株高、分蘗數及葉片數等指標的影響均不明顯,對塊莖的影響則較顯著,表現為塊莖數量與重量顯著下降。
2、低氧對苦草根系的影響顯著,表現為根須變細且數量增加,根系活力明顯下降。
3、低氧處理使岸邊沉積物處理組的沉積物氧化還原電位顯著下降、水體營養鹽濃度上升,尤其是磷酸鹽濃度顯著增加。低氧對苦草生長的影響雖不明顯,但對其種群擴張有潛在的不利作用。
.jpg)
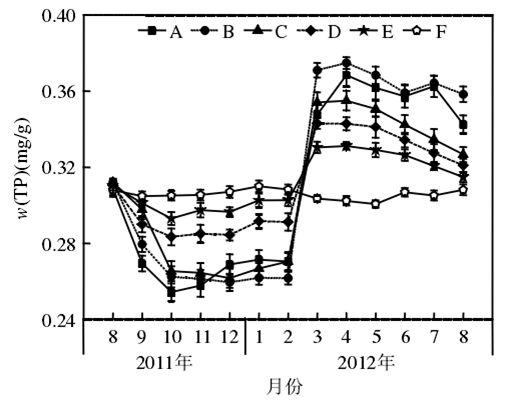
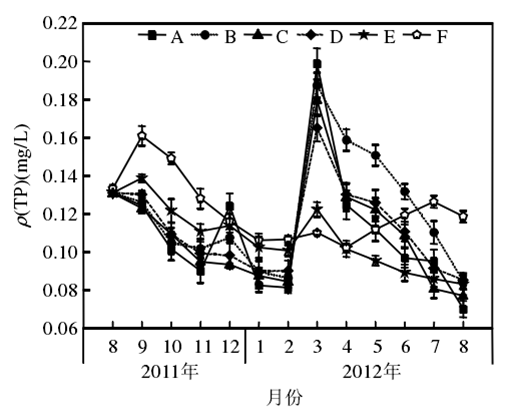
5.2 水體的營養水平對苦草生長的影響
1、隨著營養鹽含量的升高,苦草生物量的增長率逐漸降低,當水中ρ(TN) 達到10 mg/L和ρ(TP) 達到0.4 mg/L時,苦草的生物量開始減少;營養鹽含量升高對苦草葉片特征的影響不明顯,而苦草根狀莖的生物量卻隨著營養鹽含量的升高而逐漸減少。
2、當水體營養水平達到ρ(TN) 為10 mg/L和ρ(TP) 為0.4 mg/L時,除葉片長度外,苦草的其他形態指標值均顯著下降。
3、當水中磷含量增加1 倍時對苦草生長造成的抑制作用大于氮含量增加1倍時;當二者均增加時,對植物生長造成的抑制作用顯著增加。
5.3 水深光照的影響
1、苦草的水深分布
洱海苦草定植的水深范圍為 0.5~5.6 m,撫仙湖苦草生長的最大水深為20m。鄱陽湖水淹深度7.2m 以內時冬芽萌發率和水深相關不密切, 超過 7.2m 以上時呈明顯的負相關。
.jpg)
苦草分布湖泊的水質狀況
2、光照需求范圍
1)苦草幼苗正常生長發育所需的最小相對光照強度為1.6%。
2)弱光照和高營養均能對苦草生長產生抑制,兩者有交互作用,主要是弱光影響高營養的抑制作用。水下光照低于10%水面光照后,弱光照成為苦草生長的主要限制因子。
3)水下光強不足自然光強的4.5%,幼苗葉片出現發黃、折斷現象,幼苗光合作用受到明顯抑制。水下的光強為水面光強的43.3%以上,苦草幼苗也受到抑制,光強范圍大約是7.1%~43.3%。
(1).jpg)
.jpg)
5.4 水流對苦草的影響
1、苦草在靜水水體中普遍存在,其強大的根系和柔軟的帶狀葉使它能很好地生活在流動的水體中,即使在流速較大的溪流生境中也能生存。野外調查發現流速達到 0. 5 m•s- 1以上的生境中都能很好地生存。
2、為了更好的了解水流對苦草的影響,有研究人員層用水槽進行模擬流水,得出以下結論:
(1) 冠層以上,水流不受苦草冠層的影響,時均流速呈對數分布; 冠層內部,由于冠層阻流面積在垂線分布上存在差異,冠層內時均流速呈逆梯度分布,且在床面附近出現局部流速最大值。
(2) 冠頂處,水流強烈的紊動摻混與流速切變使得雷諾應力在冠頂處達到最大值,并向水面與床底方向逐漸減小; 冠層內部,逆流速梯度的出現使得雷諾應力出現負值,并在冠層內部出現局部最大值。
(3) 切應力產生的剪切紊動使得紊動能在冠頂附近最大,并向冠層內部進行垂向傳輸。受傳輸距離的限制,冠層底部水流以尾流紊動為主,紊動能大大減小。
.jpg)
5.5 植物化感與矮化處理
1、苦草可分泌化感物質酚酸,主要組分為:苯甲酸、對羥基苯甲酸、對羥基苯乙酸、鄰苯二甲酸、對羥基苯丙酸、香草酸、原兒茶酸、阿魏酸和咖啡酸等九種酚酸。 酚酸抑藻測試的結果顯示其抑藻活性與本身的結構有關,不同酚酸以毒性效應比例多維混合表現出加和抑藻效應,且隨著混合種數的增多,酚酸的加和效應增強。
2、矮壯素2-氯乙基三甲基氯化銨濃度為 0. 02 g•L- 1左右時,對苦草矮化較為適宜。而矮壯素對自然條件下深水湖泊和開放性水體的生態影響,有待進一步研究。
六、思考與展望
1、目前關于苦草的研究主要集中在實驗室階段和野外調研,關于苦草的應用研究相對較少。
2、關于苦草的研究缺乏系統性和深度的研究,目前仍有較大的不確定性。
3、工程應用問題的解決必須建立在已有的基礎之上,靈活運用自然規律和科學方法。盲目的進行開展工作,不僅會帶來極大的資源浪費,還有可能徒勞無獲。
參考文獻:
[1]劉曉培,張飲江,李 巖,等.矮壯素對苦草矮化特征及生理指標的影響[J].生態學雜志 2012,31( 10) : 2561-2567.
[2]沈顯生.安徽苦草屬一新種[J].植物分類學報(2001)39(6):571~ 574.
[3]沈顯生.安徽省苦草屬種子的掃描電鏡觀察[J].暨南大學學報2001,22 (1 ):89-91.
[4]王春景 , 周守標 , 楊海軍,等.安徽水生維管植物的多樣性[J]南京林業大學學報2006,30(5)87-90.
[5]朱增銀,尹大強,陳燦,等.不同比例硝態氮和尿素氮對苦草的生理影響 [J]南京大學學報(自然科學)2005,41(6):627-633.
[6]黃玉源,雷澤湘,何柳靜,等. 不同磷濃度對水生植物苦草的影響研究[J]. 環境科學與技術, 2011, 34(2):12- 16.
[7]宋睿,姜錦林,耿金菊,等.不同濃度銨態氮對苦草的生理影響中國環境科學[J].2011,31(3):448~453
[8] 中國植物志編委會.中國植物志(第8 卷)[ M] .北京:科學出版社, 1992.176 ~ 181.


